
 journal menu
journal menu









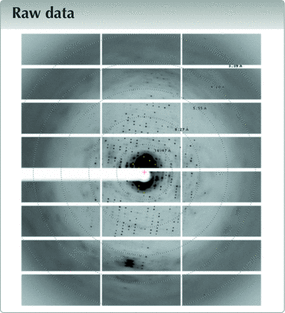
raw data letters
 | IUCrDATA |
 access
accessX-ray diffraction images for two membrane protein crystals presenting high anisotropy; the B. subtilis ABC transporter BmrA and the S. pneumoniae NADPH oxidase

aEMBL Grenoble, 71 Avenue des Martyrs, Grenoble, France, bUniv. Grenoble Alpes, CNRS, CEA, Institut de Biologie Structurale, Grenoble, France, cInstitut Universitaire de France, Paris, France, and dMolecular Microbiology & Structural Biochemistry, UMR5086, CNRS University Lyon-1, France
*Correspondence e-mail: [email protected]
Keywords: membrane protein; diffraction anisotropy.

Metadata imgCIF file: https://doi.org/10.1107/S2414314625005917/he4560img1.cif
Metadata imgCIF file: https://doi.org/10.1107/S2414314625005917/he4560img2.cif
Metadata imgCIF file: https://doi.org/10.1107/S2414314625005917/he4560img3.cif
Introduction
Diffraction anisotropy is a phenomenon more prevalently encountered with membrane protein crystals (Robert et al., 2017![]() ), which results in artefacts in electron density maps making them difficult to interpret and prone to error in model building. Our previous study suggests that anisotropy would be linked to collinearity of secondary structure elements that cause stronger diffraction peaks in the 5 Å resolution limit area (Martin et al., 2021
), which results in artefacts in electron density maps making them difficult to interpret and prone to error in model building. Our previous study suggests that anisotropy would be linked to collinearity of secondary structure elements that cause stronger diffraction peaks in the 5 Å resolution limit area (Martin et al., 2021![]() ). This collinearity of secondary structure elements is always observed in membrane proteins where trans-membrane segments are normal to the membrane plane. The data presented here (Table 1
). This collinearity of secondary structure elements is always observed in membrane proteins where trans-membrane segments are normal to the membrane plane. The data presented here (Table 1![]() ) are for two membrane proteins showing typical low resolution and strong anisotropy that represented a major challenge in their structure solving. It is believed that the diffraction anisotropy present in the data is in fact the result of two unrelated phenomena. There is (i) a difference in resolution limits linked to the crystal packing that complicates the definition of a single resolution limit for the dataset, and (ii) a difference in intensity falloff in 3D compared to the empirical curve first described by Popov & Bourenkov (2003
) are for two membrane proteins showing typical low resolution and strong anisotropy that represented a major challenge in their structure solving. It is believed that the diffraction anisotropy present in the data is in fact the result of two unrelated phenomena. There is (i) a difference in resolution limits linked to the crystal packing that complicates the definition of a single resolution limit for the dataset, and (ii) a difference in intensity falloff in 3D compared to the empirical curve first described by Popov & Bourenkov (2003![]() ), leading to the idea that membrane proteins don't follow the empirical law derived from highly diffracting crystals of soluble proteins. The data presented here can be used by methods developers to test various hypotheses on how to handle such datasets.
), leading to the idea that membrane proteins don't follow the empirical law derived from highly diffracting crystals of soluble proteins. The data presented here can be used by methods developers to test various hypotheses on how to handle such datasets.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The B. subtilis multidrug ABC transporter BmrA
The inactive catalytic mutant E504A was used to trap the transporter in the outward-facing conformation in complex with ATP-Mg2+. The transporter is a homodimer and crystallized in the P21 with two homodimers in the The solvent content is around 69%, typical for a membrane protein crystallized in detergent. The crystal that gave the highest resolution data was a large, long rod (Fig. 1![]() ). The crystal was illuminated using a microfocus beam on beamline PX2 (Duran et al., 2013
). The crystal was illuminated using a microfocus beam on beamline PX2 (Duran et al., 2013![]() ) at SOLEIL using two collection strategies. First it was collected at a single spot at 5% transmission for a low-dose data collection. This data set is included in the Zenodo deposition named ‘V-CK63-8-ld_1_data_xxxxxx.h5'. Second, diffraction data were collected with a helical crystal translation strategy at nearly 100% transmission after moving the crystal so that the beam was hitting a different (previously unexposed) part of the crystal compared to the previous datasets; this data set is included in the Zenodo deposition named ‘V-CK63-8-helical_1_data_xxxxxx.h5'. For information, by using a microfocus beam on such a large crystal we hoped to identify subdomains of better diffracting quality. Unfortunately, we were not able to identify any as judged by visual inspection of the diffraction patterns.
) at SOLEIL using two collection strategies. First it was collected at a single spot at 5% transmission for a low-dose data collection. This data set is included in the Zenodo deposition named ‘V-CK63-8-ld_1_data_xxxxxx.h5'. Second, diffraction data were collected with a helical crystal translation strategy at nearly 100% transmission after moving the crystal so that the beam was hitting a different (previously unexposed) part of the crystal compared to the previous datasets; this data set is included in the Zenodo deposition named ‘V-CK63-8-helical_1_data_xxxxxx.h5'. For information, by using a microfocus beam on such a large crystal we hoped to identify subdomains of better diffracting quality. Unfortunately, we were not able to identify any as judged by visual inspection of the diffraction patterns.
![[Figure 1]](he4560fig1thm.gif) | Figure 1 ABC transporter BmrA. (a) A crystal in a hanging drop. The diffracting crystal is in the cyan rectangle. (b) The same crystal viewed from the PX2 beamline camera, probably before helicoidal data collection. (c) The crystal packing and unit cell. The asymmetric unit is shown in blue. (d) Typical diffraction image. |
The structure of BmrA solved and refined using only the helical crystal translation data from this crystal has been published as part of the article by Chaptal et al. (2022![]() ), with a detailed material and methods section on structure solving and model building, which were challenging for this project.
), with a detailed material and methods section on structure solving and model building, which were challenging for this project.
The S. pneumoniae NADPH oxidase
A F397W mutant of the S. pneumoniae NADPH oxidase (SpNOX) was crystallized with the trans membrane domain complexing two hemes and the dehydrogenase (DH) domain complexed with FAD. The full-length F397W SpNOX in detergent was crystallized as a monomer in P6422 with a high solvent content of 76%. The typical crystal shape was a nice hexagonal firebrick (Fig. 2![]() ). More than 500 crystals were exposed to X-rays on the MASSIF-1 automatic beamline (Bowler et al., 2015
). More than 500 crystals were exposed to X-rays on the MASSIF-1 automatic beamline (Bowler et al., 2015![]() ) at ESRF, Grenoble and 187 diffraction datasets were recorded and processed; among them only 32 reached resolutions better than 4 Å. A high-resolution structure is also available for the DH domain alone (https://doi.org/10.2210/pdb8qq5/pdb) (Petit-Hartlein et al., 2024
) at ESRF, Grenoble and 187 diffraction datasets were recorded and processed; among them only 32 reached resolutions better than 4 Å. A high-resolution structure is also available for the DH domain alone (https://doi.org/10.2210/pdb8qq5/pdb) (Petit-Hartlein et al., 2024![]() ) that was used to solve our structure using molecular replacement. Two search models were used, first the DH domain was placed, then a model of the transmembrane domain made on the basis of multiple sequence alignment of NOX homologs and validated by structural alignment on CsNOX, another procaryotic NOX (Magnani et al., 2017
) that was used to solve our structure using molecular replacement. Two search models were used, first the DH domain was placed, then a model of the transmembrane domain made on the basis of multiple sequence alignment of NOX homologs and validated by structural alignment on CsNOX, another procaryotic NOX (Magnani et al., 2017![]() ). This high-resolution structure of the DH domain was also used during our model refinement as a reference template during model building. For the final refinement we used software dedicated to low resolution structures modelling, ISOLDE (Croll, 2018
). This high-resolution structure of the DH domain was also used during our model refinement as a reference template during model building. For the final refinement we used software dedicated to low resolution structures modelling, ISOLDE (Croll, 2018![]() ) and LORESTR (Kovalevskiy et al., 2016
) and LORESTR (Kovalevskiy et al., 2016![]() ) as described in the original paper (Petit-Hartlein et al., 2024
) as described in the original paper (Petit-Hartlein et al., 2024![]() ).
).
| | Figure 2 NADPH oxidase SpNOX. (a) A typical spNOX crystal in hanging drops. (b) Crystal packing and unit cell for SpNOX. The asymmetric unit is shown in blue. (c) Typical diffraction image. |
The structure of SpNOX solved and refined using data from this crystal was published as part of this article (Petit-Hartlein et al., 2024![]() ), with a detailed material and methods section on structure solving and model building, which as mentioned above was challenging for this project.
), with a detailed material and methods section on structure solving and model building, which as mentioned above was challenging for this project.
Supporting information
Link https://doi.org/10.5281/zenodo.14901515
DOI for imageCIF 8QQ7.cif
Metadata imgCIF file. DOI: https://doi.org/10.1107/S2414314625005917/he4560img1.cif
Metadata imgCIF file. DOI: https://doi.org/10.1107/S2414314625005917/he4560img2.cif
Metadata imgCIF file. DOI: https://doi.org/10.1107/S2414314625005917/he4560img3.cif
Funding information
This work was supported by the Centre National de la Recherche Scientifique (CNRS), Lyon University and the Grenoble-Alpes University. VZ was supported by the Lyon-1 university EDISS doctoral school. This work was also supported by the French Agence Nationale de la Recherche Projects ANR-19-CE11-0023–01 for VZ, PF and VC, and ANR17-CE11-0013 to FF. The BmrA project benefitted from SFR Biosciences (Universite Claude Bernard Lyon 1, CNRS UAR3444, Inserm US8, ENS de Lyon) Protein Science Facility with the help of Virginie Gueguen-Chaignon, especially. The SpNOX work used the platforms of the Grenoble Instruct center (UMS 3518 CNRS-CEA-UJF-EMBL) with support from FRISBI (ANR-10- INSB-0502) and GRAL (ANR10-LABX-49–01) within the Grenoble Partnership for Structural Biology. AV was supported through the Emergence program from the University Grenoble Alpes.
References
![]() Bowler, M. W., Nurizzo, D., Barrett, R., Beteva, A., Bodin, M., Caserotto, H., Delagenière, S., Dobias, F., Flot, D., Giraud, T., Guichard, N., Guijarro, M., Lentini, M., Leonard, G. A., McSweeney, S., Oskarsson, M., Schmidt, W., Snigirev, A., von Stetten, D., Surr, J., Svensson, O., Theveneau, P. & Mueller-Dieckmann, C. (2015). J. Synchrotron Rad. 22, 1540–1547. Web of Science CrossRef CAS IUCr Journals Google Scholar
Bowler, M. W., Nurizzo, D., Barrett, R., Beteva, A., Bodin, M., Caserotto, H., Delagenière, S., Dobias, F., Flot, D., Giraud, T., Guichard, N., Guijarro, M., Lentini, M., Leonard, G. A., McSweeney, S., Oskarsson, M., Schmidt, W., Snigirev, A., von Stetten, D., Surr, J., Svensson, O., Theveneau, P. & Mueller-Dieckmann, C. (2015). J. Synchrotron Rad. 22, 1540–1547. Web of Science CrossRef CAS IUCr Journals Google Scholar
![]() Chaptal, V., Zampieri, V., Wiseman, B., Orelle, C., Martin, J., Nguyen, K. A., Gobet, A., Di Cesare, M., Magnard, S., Javed, W., Eid, J., Kilburg, A., Peuchmaur, M., Marcoux, J., Monticelli, L., Hogbom, M., Schoehn, G., Jault, J. M., Boumendjel, A. & Falson, P. (2022). Sci. Adv. eabg9215. Google Scholar
Chaptal, V., Zampieri, V., Wiseman, B., Orelle, C., Martin, J., Nguyen, K. A., Gobet, A., Di Cesare, M., Magnard, S., Javed, W., Eid, J., Kilburg, A., Peuchmaur, M., Marcoux, J., Monticelli, L., Hogbom, M., Schoehn, G., Jault, J. M., Boumendjel, A. & Falson, P. (2022). Sci. Adv. eabg9215. Google Scholar
![]() Croll, T. I. (2018). Acta Cryst. D74, 519–530. Web of Science CrossRef IUCr Journals Google Scholar
Croll, T. I. (2018). Acta Cryst. D74, 519–530. Web of Science CrossRef IUCr Journals Google Scholar
![]() Duran, D., Couster, S. L., Desjardins, K., Delmotte, A., Fox, G., Meijers, R., Moreno, T., Savko, M. & Shepard, W. (2013). J. Phys. Conf. Ser. 425, 012005. CrossRef Google Scholar
Duran, D., Couster, S. L., Desjardins, K., Delmotte, A., Fox, G., Meijers, R., Moreno, T., Savko, M. & Shepard, W. (2013). J. Phys. Conf. Ser. 425, 012005. CrossRef Google Scholar
![]() Kovalevskiy, O., Nicholls, R. A. & Murshudov, G. N. (2016). Acta Cryst. D72, 1149–1161. Web of Science CrossRef IUCr Journals Google Scholar
Kovalevskiy, O., Nicholls, R. A. & Murshudov, G. N. (2016). Acta Cryst. D72, 1149–1161. Web of Science CrossRef IUCr Journals Google Scholar
![]() Magnani, F., Nenci, S., Millana Fananas, E., Ceccon, M., Romero, E., Fraaije, M. W. & Mattevi, A. (2017). Proc. Natl Acad. Sci. USA 114, 6764–6769. Web of Science CrossRef CAS PubMed Google Scholar
Magnani, F., Nenci, S., Millana Fananas, E., Ceccon, M., Romero, E., Fraaije, M. W. & Mattevi, A. (2017). Proc. Natl Acad. Sci. USA 114, 6764–6769. Web of Science CrossRef CAS PubMed Google Scholar
![]() Martin, J., Robert, X., Gouet, P., Falson, P. & Chaptal, V. (2021). bioRxiv 2021.2012.2007.471609. Google Scholar
Martin, J., Robert, X., Gouet, P., Falson, P. & Chaptal, V. (2021). bioRxiv 2021.2012.2007.471609. Google Scholar
![]() Petit-Hartlein, I., Vermot, A., Thepaut, M., Humm, A.-S., Dupeux, F., Dupuy, J., Chaptal, V., Marquez, J. A., Smith, S. M. E. & Fieschi, F. (2024). eLife RP93759. Google Scholar
Petit-Hartlein, I., Vermot, A., Thepaut, M., Humm, A.-S., Dupeux, F., Dupuy, J., Chaptal, V., Marquez, J. A., Smith, S. M. E. & Fieschi, F. (2024). eLife RP93759. Google Scholar
![]() Popov, A. N. & Bourenkov, G. P. (2003). Acta Cryst. D59, 1145–1153. Web of Science CrossRef CAS IUCr Journals Google Scholar
Popov, A. N. & Bourenkov, G. P. (2003). Acta Cryst. D59, 1145–1153. Web of Science CrossRef CAS IUCr Journals Google Scholar
![]() Robert, X., Kassis-Sahyoun, J., Ceres, N., Martin, J., Sawaya, M. R., Read, R. J., Gouet, P., Falson, P. & Chaptal, V. (2017). Sci. Rep. 7, 17013. Web of Science CrossRef PubMed Google Scholar
Robert, X., Kassis-Sahyoun, J., Ceres, N., Martin, J., Sawaya, M. R., Read, R. J., Gouet, P., Falson, P. & Chaptal, V. (2017). Sci. Rep. 7, 17013. Web of Science CrossRef PubMed Google Scholar
This is an open-access article distributed under the terms of the Creative Commons Attribution (CC-BY) Licence, which permits unrestricted use, distribution, and reproduction in any medium, provided the original authors and source are cited.
| IUCrDATA |



